|
Введение в моделирование систем |
|
| |
|
Модели биосистем |
|
Введение в моделирование систем |
|
| |
|
Модели биосистем |
|
Стадия разработки! >> Приглашаю Стажера-Студента для оформления HTML-страниц! \ Помогу чем могу! | |
|
| |
|
План главы |
Литература 1. Емельянов-Ярославский Л.Б. Интеллектуальная квазибиологическая система. Индуктивный автомат --> 2. Теория функциональных систем П.К.Анохина 3. Косяков Ю.Б. Мой мозг. Строение, принципы работы, моделирование. 4. Цыганков В.Д. Нейрокомпьютер и мозг. 5. Жданов А.А. The mathematical models of neuron and neural network in autonomous adaptive control methodology 6. Билич Г., Назарова Л. Популярная медицинская энциклопедия. М.: Вече. - 2004. - 384 с. 7. Лурия А.Р. Основы нейропсихологии. М.: ИЦ Академия, 2004- 384 с. 8. |
|
ОБЗОР ПОДХОДОВ К ПОСТРОЕНИЮ МОДЕЛЕЙ Вводные замечания Теория функциональной системы Анохина П.К. Автономная адаптивная система Жданова А.А. Интеллектуальная квазибиологическая система Л.Б.Емельянова-Ярославского ОБЪЕКТ ИССЛЕДОВАНИЯ - МОЗГ Нейрон Количественный и качественные характеристики \ примеры Математическая модель «хищник-жертва» (модель Вольтерра) Заключительные замечания и предложения | |
|
Заключение | Предисловие | Глоссарий | Тесты |
|
Модели биосистем
Вводные замечания
Метод познания путем моделирования -
часть так называемого системного
подхода,
то есть разумного подхода к исследованию
систем.
Самый сложный живой биологический объект с точки зрения кибернетики - безусловно - человеческий мозг.
Подходы, наработанные для моделирования технических и социальных систем в рамках теории систем, применительно к биологическим систем, особенно такому сложному, как мозг, применимы и дают некоторые результаты.
Пытаясь применить подход "разделяй и
властвуй", мозг можно рассматривать на трех уровнях:
- биофизическом или клеточном;
- нейрофизиологическом, мозг как
структура;
- психическом и
психологическом, или сознание, подсознание, сверхсознание.
На каждом уровне – своя иерархия моделей.
Работы по созданию интеллектуальных систем Емельянов-Ярославский в (__) разделял на информационное и биологическое направления.
У сторонников первого - информационного направления есть реальные действующие макеты и программы, моделирующие отдельные стороны интеллекта.
Большинство специалистов в области искусственного интеллекта исходя из того что их системы не должны повторять в своей структуре и функционировании структуру и процессы, присущие биологическим системам.
Результатов у второго направления "меньше, чем надежд".
Подход Емельянова-Ярославского можно рассматривать как реализацию познавательной стороны моделирования - возможности заглянуть в суть изучаемых явлений, лучше понять их.
ОБЗОР ПОДХОДОВ К ПОСТРОЕНИЮ МОДЕЛЕЙ
Теория функциональной системы П.К. Анохина
Исходной посылкой теории функциональной системы П.К. Анохина служит: "Цель всегда опережает реализацию ее организмом". Живая высокоорганизованная система формирует потребности в получении результата (модель цели в классическом системном подходе) или сама ставит себе цель уже в самом начале поведенческого акта.
В рамках теории расширено понятие система.
В классическом механистическом толковании для объектов исследования, допускающих расчленение на части - элементы, описание системы допускается как совокупности моделей (по Сафонову) структуры, поведения во времени, цели и ресурсов функционирования.
По П.К. Анохину "системой можно назвать только такой комплекс избирательно вовлеченных компонентов, у которых взаимодействие и взаимоотношение приобретает характер взаимосодействия компонентов на получение фокусированного полезного результата".
Основные положения теории.
Нейрон в нервной системе имеет многочисленные потенциально возможные степени свободы (режимы возбуждения и связи). Конечный результат выступает системообразующим фактором, который освобождает части большой системы от избыточных степеней свободы, то есть упорядочивают систему.
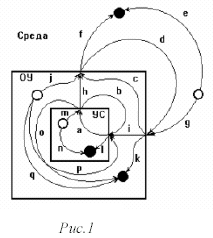
Схема функциональной системы приведена на рис. ___
Состав системы обычно идентифицируют либо по анатомическому типу или по типу функционирования. Однако в целом организме ни один полноценный результат нельзя получить только за счет одной анатомической системы: кровообращения, мышечной, дыхательной, а только при их взаимосодействии. Речь идет о системе как гетерогенном образовании.
Приняты следующие обозначения: А- стадия афферентного синтеза, ОР - ориентировочный рефлекс, ОА - обстановочная афференция, В - принятие решения, С - формирование акцептора действия и эфферентной программы самого действия, Д - получение полезного результата действия и Е - формирование обратной (санкционирующей) афференции для сличения полученного результата с запрограммированным.
Концепция "автономного адаптивного управления" как модель нервной системы
Излагается по А.А. Жданов: "Метод автономного адаптивного управления , его свойства и приложения". М.: Перспективные информационные системы и интеллектуальные системы. стр.4-14
"Подавляющее большинство автоматически управляемых технических систем, разработанных в прошедшем ХХ веке, обязаны своим существованием теории управления, построенной на аналитическом понимании законов механики и физики. Этот «классический» подход к управлению строится на том, что положение точки (объекта управления) в пространстве признаков известно абсолютно. Далее формальные математические преобразования позволяют получить математическую зависимость входов и выходов для системы управления (УС). Однако при всей изощренности наработанного математического инструментария, областью применения «классических» методов управления остаются сравнительно простые объекты управления с очевидными свойствами. Попытки аналитически описать более тонкие свойства объектов управления (технических и, тем более, естественных), особенно в случаях, когда свойства ОУ плохо определены априори, быстро приводят к катастрофическому усложнению математических моделей. Ситуацию в целом не спасают ни эвристические приемы, ни повышение эффективности вычислительной техники. На практике объекты управления, которые плохо формализуются, свойства которых априори плохо известны или изменяются в процессе функционирования, являются типичными. С середины ХХ века активно развивается «неклассический» подход в теории управления. Такие «неклассические» методы управления видят ОУ не как абсолютно известную точку в пространстве признаков, а лишь как некоторую информацию об этой «точке» (согласно А.В. Чечкину). Управление при этом сводится к формальной работе с этой информацией. Аналитические функциональные зависимости параметров заменяются априорными, либо эмпирическими знаниями, либо результатами обучения на примерах."
При разработке модели нервной системы Автор отталкивался от наблюдаемых свойств нервной системы.
|
|
1. Условие автономности - требует рассматривать управляющую систему (УС) как подсистему объекта управления (ОУ) развивающуюся вместе с ним, и самостоятельно добывающую знания, необходимые для управления.
2. Условие дискретности - строение нервной системы предполагается дискретным. Нейроны, нервные волокна, нервные импульсы -
3. Условие максимальной начальной приспособленности отражает отражает невозможность создания УС, обладающей способностью адаптироваться к любым предъявляемым объектам управления и внешней среде.
4. Условие минимума начальных знаний отражает тот факт, что нервная система новорожденного организма обладает относительным минимумом знаний и должна в течение жизни накапливать знания , необходимые для управления, то есть быть адаптивной управляющей системой. |
|
|
Целевыми функциями нервной системы полагается выживание организма и стремление к накоплению знаний. Исходя из условий и целевых функций управляющая система должна решать взаимосвязанные задачи, а именно: - находить во входных данных неслучайную, закономерную информацию - формировать образы; - распознавать в текущей входной информации сформированные образы - распознавать их; - обнаруживать специальные образы - знания, отражающие неслучайные причинно-следственные связи событий в системе, которые можно использовать для управления; - выводить новые знания из уже имеющихся; - вырабатывать эмоциональные оценки для сформированных образов; - принимать решения в каждый текущий момент времени. |
Интеллектуальная квазибиологическая система Л.Б.Емельянова-Ярославского
Нейронно-энергетическая концепция индуктивного автомата (см. www.aha.ru/~pvad/f0.htm) состоит в следующем.
Индуктивный автомат состоит из нейронов, рецепторов, эффекторов и системы раздачи питания нейронам. Эти, как и другие названия, условны. Так нейроны автомата весьма отдаленно напоминают живые нейроны. рецепторы воспринимают воздействия внешней среды, в которой находится автомат, и преобразуют их воздействия на нейроны. Эффекторы воспринимают воздействия нейронов и преобразуют их в воздействия на внешнюю среду. Под внешней средой понимается реальный мир или искусственная среда, обладающая закономерными свойствами.
Основные свойства нейронов следующие.
1. Нейрон существует в условиях метаболического обмена со средой, т.е. нуждается в питании. Если нейрон не получает требуемого ему количества питания, то он погибает.
2. Нейрон стареет.
3. В нейроне возможны функциональные обновления - особые события, приводящие к снижению возраста, а значит, и к снижению потребностей в питании.
4. Между нейронами могут образовываться связи, по которым осуществляется взаимодействие нейронов
5. Разряд в нейроне требует дополнительных расходов питания, зависящих от режима разрядов - от частоты генерации разрядов.
6. У нейрона имеются собственные внутренние резервы питания
7. Помимо контактного взаимодействия на основе возбуждающих и тормозных связей, между нейронами имеет место полевое взаимодействие, обусловленное тем, что самочувствие одних нейронов изменяет некоторые характеристики у других нейронов.
Автомат в исходном состоянии - это неорганизованное или слабоорганизованное множество нейронов, поставленных в условия ограниченного питания.
Нейрон - основной элемент нервной системы - клетка через которую происходит передача и преобразование информации.
Нервная клетка состоит из тела клетки (называемого также сомой) и отходящих от него отростков: коротких входных дендритов и длинного аксона, по которому информация передается от одного нейрона к другому.
|
Рис. 1 |
Синапс на ядре Синапс на дендрите Аксон Ядро Дендриты Ядро следующей клетки |
Область, в которой возбуждение от одного нейрона передается к другому, называется синаптическим переходом или синапсом. В соединениях между нейронами участвует либо само тело клетки, либо его тонкие отростки - дендриты. Аксон оканчивается либо небольшим числом ветвей, либо таких ветвей много, и тогда на выходе одного нейрона образуется множество синаптических соединений с другими нейронами.
Сходным образом тело одного нейрона может принимать сигналы либо от нескольких, либо от тысяч других нейронов.
Нервные волокна и дендриты, с точки зрения Технаря, можно рассматривать как изолированные проводники, по которым передаются сигналы, характерные для нервной системы - электрические импульсы. Эти электрические, а точнее, ионные сигналы генерируются в клеточном теле в ответ на активность синаптических входов. В самом синапсе под влиянием поступающего импульса высвобождается особое химическое вещество - медиатор; оно пересекает зазор между окончанием аксона и рецепторным участком следующего нейрона и вызывает изменение нормального электрического потенциала данного нейрона. Если количество медиатора достаточно велико, изменение электрического потенциала нейрона достигает предела, при превышении которого рождается импульс; затем импульс идет по аксону, пока не достигнет следующего синаптического соединения, где весь процесс повторяется.
Лишь в редких случаях активности в одном синапсе достаточно для появления нового импульса. Как правило, нейрон должен получить много импульсов, прежде чем в нем возникнет ответный импульс.
Все синаптические соединения делятся на два класса: возбудительные и тормозные. Активность первых увеличивает вероятность ответа нейрона, активность вторых - уменьшает.
Ответ нейрона на активность всех его синапсов представляет собой как бы результат “химического голосования”: соотношение “голосов” возбудительных и тормозных синапсов на входе нейрона определяет уровень его активности. Частота ответов нейрона в общем случае зависит от частоты и характера стимулов на его синапсах. Но имеются ограничения. Генерация импульса делает нейрон недееспособным в течении 0,001 секунды. То есть максимальная теоретическая частота ответа нейрона равна 1000 импульсам в секунду. Практически она еще ниже от 300 до 800 импульсов в секунду.
<Тиганов А.С.>
Цитоархитектоника головного мозга человека организована таким образом, что более чем 10 млрд. нервных клеток, занимая относительно небольшое пространство и будучи сформированными в специализированные структуры, обеспечивают специфические функции мозга, связанные с восприятием, переработкой и проведением информации, в соответствии с которой осуществляется взаимодействие организма с внешней средой на основе высокой нейрональной специфичности и пластичности.
Основной структурной единицей нервной системы является нейрон. Различные типы нейронов дифференцируются по величине и форме тела клетки, а также по длине и степени ветвистости ее отростков.
Рецепторы нейронов — это белковые структуры, расположенные на внешней поверхности мембраны клеток. Они способны "распознавать" и связывать биологически активные вещества — нейротрансмиттеры, различные эндогенные вещества, а также экзогенные соединения, в том числе психофармакологические средства. Соединения, которые могут связывать рецепторы, называются лигандами. Лиганды бывают эндогенными и экзогенными.
Распознавание лиганда рецептором обеспечивается специальными структурными элементами, или сайтами. Специфичность связывания лиганда происходит благодаря структурному соответствию молекул лиганда и рецептора, когда они подходят друг к другу по типу "ключ к замку". Реакция связывания является моментом запуска каскада внутриклеточных реакций, приводящих к изменению функционального состояния нейрона. В зависимости от "силы" и "прочности" связывания лиганда с рецептором употребляют понятие аффинности (сродства) лиганда по отношению к рецептору.
При связывании рецептора с лигандом может происходить как активация, так и блокада рецептора. В связи с этим говорят об агонистах и антагонистах рецепторов, а также о частичных агонистах (рис. 7).
Максимальную эффективность в отношении активации рецептора имеет полный агонист, минимальную (практически нулевую) — антагонист. Между ними находятся вещества, называемые частичными агонистами. Последние действуют значительно мягче, чем полные агонисты. Частичные агонисты, кроме того, занимая определенное пространственное положение в молекуле рецептора, могут предотвращать избыточное действие полного агониста, т.е. действуют частично как антагонисты. В этом случае употребляют понятие агонист/антагонист.
Высокой аффинностью могут обладать как агонисты, так и антагонисты рецептора. Агонист активирует рецептор, вызывая соответствующий физиологический эффект, в то время как антагонист, связываясь с рецептором, блокирует его и предотвращает развитие физиологического эффекта, выявляемого агонистами. Примером антагонистов могут служить нейролептики, которые предотвращают эффекты дофамина на уровне дофаминового рецептора.
При связывании лиганда с рецептором происходит изменение конфигурации последнего (рис. __).
Многие вещества, как эндогенные, так и экзогенные, реагируют не с одним, а с несколькими типами рецепторов — "семейством" их, которое подразделяется на отдельные типы. Примером могут служить многие нейротрансмиттеры, реагирующие с несколькими типами специфических рецепторов (например, Д1—Д5-типы дофаминовых рецепторов). Существование нескольких рецепторов к одному лиганду носит название гетерогенности рецепторов.
Представление о функции рецепторов было бы неполным, если не представить внутриклеточные процессы, развивающиеся после связывания рецептора соответствующим веществом, и механизмы, обеспечивающие трансформацию внешнего сигнала в процессы, приводящие к появлению нервного импульса. Связывание лиганда с рецептором может приводить либо непосредственно к открытию (или закрытию) соответствующих ионных каналов (см. рис. __), либо к активации вторичных мессенджерных систем (в качестве первичного мессенджера рассматривается вещество, реагирующее с рецептором).
Первые упоминания о вторичных мессенджерных системах появились в связи с работами E.Sutherland и соавт. (1950), которые показали, что адреналин стимулирует гликогенез путем увеличения концентрации циклического аденозинмонофосфата (цАМФ) в клетке. Оказалось, что этот вторичный мессенджер опосредует и другие клеточные реакции. В дальнейшем была выявлена связь действия цАМФ с активацией белковых киназ — ферментов, фосфорилирующих белки, что приводит к изменению их структуры и активности.
Позднее были открыты и другие вторичные мессенджеры. Сейчас выделяют среди них 3 класса: 1) циклические нуклеотиды (цАМФ, циклический гуанозинмонофосфат — цГМФ); 2) ионы кальция (Са2+); 3) метаболиты фосфолипидов — инозитол-1,4,5-трифосфат (1Р3), диглицерин (ДАГ), арахидоновую кислоту. В отличие от других вторичных мессенджеров Са2+ транспортируется в нейрон из внутриклеточного пространства.
Мембраны нейрона содержат специализированные трансмембранные белки, которые формируют ионные каналы не только для Са2+, но и для других ионов, концентрация которых по обе стороны мембраны влияет на изменение мембранного потенциала. Происходят поляризация и деполяризация мембраны, т.е. изменение трансмембранного потенциала. Наибольшее значение в этих процессах имеют ионные каналы для натрия (Na+), калия (К+), хлора (С1-) и кальция (Са2+).
В 60—70-х годах с помощью гистохимических и радиоиммунологических методов было установлено, что в мозге имеются дифференцированные нейрохимические системы — норадренергическая, дофаминергическая, серотонинергическая и др. Они представляют собой комплекс мозговых структур, функция которых определяется наличием общего нейротрансмиттера и рецепторов, взаимодействующих с ним. В одних из структур, входящих в нейрохимическую систему, расположены тела нейронов, в других оканчиваются терминали нервных клеток. В последнем случае говорят о проекциях на те или иные мозговые образования. В одних и тех же структурах мозга могут располагаться клетки и проекции нескольких нейрохимических систем.
Дофаминергическая система. В этой системе мозга различают 7 отдельных подсистем (систем, трактов): нигростриатную, мезокортикальную, мезолимбическую, тубероинфундибулярную, инцертогипоталамическую, диенцефалоспинальную и ретинальную. Из них первые 3 являются основными (рис. 8). Тела нейронов нигростриатной, мезокортикальной и мезолимбической систем расположены на уровне среднего мозга, образуют комплекс нейронов черной субстанции и вентрального поля покрышки. Они составляют непрерывную клеточную сеть, проекции которой частично перекрываются, поскольку аксоны этих нейронов идут вначале в составе одного крупного тракта (медиального пучка переднего мозга), а оттуда расходятся в разные мозговые структуры. Формирование нигростриатной, мезолимбической и мезокортикальной систем определяется областями, где оканчиваются аксоны дофаминергических нейронов, т.е. локализацией их проекций. Некоторые авторы объединяют мезокортикальную и мезолимбическую подсистемы в единую систему. Более обоснованным является выделение мезокортикальной и мезолимбической подсистем соответственно проекциям в лобную кору и лимбические структуры мозга (рис. __).
Нервная система содержит 10 миллиардов нервных клеток нескольких десятков типов. Зрительная система - 120 миллионов клеток первого уровня и 800 тысяч второго.
Для сравнения PENTIUM II - 7.5 миллионов транзисторов, PENTIUM IV– 42 миллиона. Прогноз развития полупроводниковой техники дает оценку 1,5 млд. транзисторов на кристалле.
Теоретически, на основе простейших нейронов несложно построить структуры, различающие границу света и тени, контур, движение, а используя элементы задержки - измерители скорости и ускорения. Пример - индикатор света/тени, построенный на суммирующем элементе с коэффициентами - 0.5, 1, -0,5.
|
5 10 10 Сигнал на выходе: = 2,5 |
Излагаемая ниже модель [___] является хорошим примером математический модели исследования системы, в данном случае популяции 2 видов животных.
Создание модели.
Описание системы. Имеется некоторая популяция 2 х видов – зайцы (жертвы) и рыси (хищника)[2], в которой происходят жизненные процессы во всем их многообразии. Зайцы питаются растительной пищей, имеющейся в достаточном количестве (между зайцами отсутствует внутривидовая борьба). Рыси могут питаться только зайцами.
Постановка задачи. Найти законы изменения численности популяции во времени.
Основные допущения.
1. Существуют только процессы размножения и естественной гибели, скорости которых пропорциональны численности особей в данный момент времени.
2. Не учитываются биохимические, физиологические процессы.
3. Нет борьбы между особями за место обитания, за пищу (бесконечно большое пространство и количество пищи.
Переменные модели:
X - число жертв в момент t,
Y – число хищников в момент t <
Уравнение баланса между численностью рожденных и гибнущих особей:
Приглашаю принять участие в обсуждении темы. Пишите -- Сазанов Владимир Михайлович -- E-mail
|
МОЗГОВЕДЕНИЕ |
Междисциплинарный семинар "МОЗГ" \ Анохин К.В. |
ГОМЕОСТАТИКА |
НейрОК
Интелсофт www.curricula.ru \ Университетские | |
|
Научно-образовательный |
Л.Б. Емельянов-Ярославский
Интеллектуальная квазибиологическая система Индуктивный автомат --> |
Степанов А.М. Основы медицинской гомеостатики. |
Горский Ю.М. Основы гомеостатики. | |
| Simulation of process of control in cardio-vascular system by means of neuron-like Autonomous Adaptive Control (AAC) system \ Жданов А.А | В.Г. Редько. Эволюционная биокибеорнетика |
Теслинов А.Г. Гармония и гомеостаз // Материалы семинара по гомеостатике. - Международный Университет, Москва, 1999. | Некоторые подходы к построению эффективных систем управления фирмами, основанные на гомеостатических принципах. \ Минзов А. С., Добренькова Е.В. | |
|
|
|
ВИРТУАЛЬНАЯ ШКОЛА КОМПЬЮТЕРНЫХ ТЕХНОЛОГИЙ |
© 2002-2005 Сазанов В.М. |
| |
| О проекте | Курсы | Условия | Экстернат | Партнерство | Семинар | Ссылки | Контакты | |||
Сегодня: